
17.1. PERFUME QUALITY AND ART
Contemporary perfumes contain from tens to hundreds of ingredients and are comprised of (1) essential oils derived from natural aromatic plant extracts and/or synthetic aromatic chemicals which are classified by structural group (e.g., alcohols, esters, aldehydes, and terpenes); (2) fixatives, natural or synthetic substances used to reduce the evaporation rate, increase perceived odor strength, and improve stability; and (3) solvents, the liquid in which the perfume oil is dissolved in. The typical solvent solution is 98% ethanol and 2% water.
The science of perfume is chemistry and the aromatic result is artistry. Indeed, especially in France, perfume creation is treated as high art. According to one French perfume executive: “…perfumers consider that what they create is great art and that because they are French the world should come on bended knee and consider itself lucky to be blessed with their creations… [they say] ‘I launched this and that perfume, and my perfumes are wonderful, fabulous, they lost five million dollars, but who cares, they’re objects of art that will live love forever and conform to my immortal, pure aesthetic’” (Burr 2007, 53–54). Perfume briefs, the perfume company’s instruction to the perfumer of what the perfume should smell (be) like, are equally artistic and vague. For example, Parfums Dior posed this brief for Pure Poison (2004): “What is it like to have something soft and hard at the same time?” (Burr 2007, 55). More typical examples are: “Give us the scent of a warm cloud floating in a fresh spring sky over Sicily raining titanium raindrops on a woman with emerald eyes” (Burr 2007, 55).
The language of perfumery bears witness to its inherently aesthetic qualities. Fragrances are designated according to their concentration level, the scent family they belong to, and the notes in the scent. The concentration level of the perfume oil in a fine fragrance indicates its intensity and its predicted duration on the skin. The more concentrated the perfume, the stronger the scent and the longer it will last. Although there is variability within the definitions, there are four major perfume concentration classifications. Parfum contains between 15% and 30% aromatic compounds; eau de parfum contains 8–15% aromatic compounds; eau de toilette ranges from 4% to 8% aromatic compounds; and eau de cologne contains between 2% and 5% aromatic compounds. Eau de cologne was originally invented by Italian perfumers living in KÖln (Cologne) Germany in the 1700s and was made from rosemary and citrus essences dissolved in wine. However, the term “cologne” has become a generic for a weakly concentrated perfume and/or a man’s fine fragrance.
The second category of perfume grouping is by scent family and scent family subtype. Scent families are designated with traditional classification terms (originating from around 1900) and modern terms (since 1945). The main scent families are Floral, Chypre, Fougère, Marine/Ozonic, Oriental, Citrus, Green, and, most recently, Gourmand. Scent family subtypes include terms such as fresh, aldehyde, amber, fruity, spicy, woody, and animalic. For further information on scent family types, see Moran (2000).
Perfumery can be likened to the nose as music is to the ear. In keeping with this aesthetic connotation, the third classification of a perfume’s olfactory quality is described in musical metaphors. The combination of ingredients in a perfume is called a “composition” and it has three “notes” that unfold over time. The first note is called the top note, or head note, and it produces the immediate impression of the perfume. Top notes consist of small, light molecules with high volatility that evaporate quickly. Middle notes (also called heart notes) emerge just before the top notes have dissipated. Scents from this note class appear anywhere from two minutes to one hour after the application of a perfume. Base notes (or bottom or dry down) appear while the middle notes are fading. Compounds of this class are often the fixatives used to hold and boost the strength of the lighter top and middle notes. Base notes are large, heavy molecules that evaporate slowly and are usually not perceived until 30 minutes after the application of the perfume. Some base notes can still be detectable 24 hours or more after application. The varying evaporation rates of different molecules in a perfume mean that a perfume will not smell the same when it is first put on as it does three hours later.
Perfume qualities are described in musical metaphors not solely because of the aesthetic relationship between perfume and music but because there are so few specific words dedicated to olfactory experience. Anthropologists have found that in all known languages, there are fewer words that refer explicitly to our experience of smells than there are for any other sensation (Classen, Howes, and Synnott 1994). In English, aromatic, fragrant, pungent, redolent, and stinky exhaust the list of adjectives that specifically describe olfactory stimuli and nothing else. More common terms used to describe odors, like floral or fruity, are references to the odor-producing objects (flowers and fruits), not the odors themselves. We also borrow terms from other senses; chocolate smells sweet, grass smells green, and so on (Herz 2005, 2008).
Various possibilities explain why our sense of smell and language are so disconnected. First, unlike other sensory systems, olfactory information does not need to be integrated in the thalamus prior to processing in the cortex, and it is argued that the thalamus has relevance for language. Second, a large body of evidence indicates that the majority of olfactory processing occurs in the right hemisphere of the brain, whereas language processing is known to be dominated by the left hemisphere (see Royet and Plailly 2004, for review). It has also been suggested that odors are hard to name because of competition between odor and language processing for cognitive resources that share the same neural substrates (Lorig 1999). This latter theory is supported by a magnetoen-cephalographic study which showed that the presence of an odor altered the semantic processing of words and degraded word encoding, but did not influence nonsemantic processing (Walla et al. 2003).
17.2. BRIEF HISTORY OF PERFUME
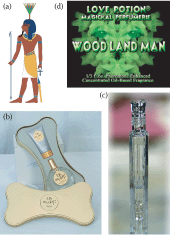
The word perfume comes from the Latin “per fumum,” meaning through smoke. It originated about 4000 years ago among the Mesopotamians in the form of incense. The original aromatic essences used in perfumery were herbs and spices like coriander and myrtle; flowers were not used until much later. Perfumery, the art of making perfume, then traveled to Egypt where it was initially only used in rituals for the gods or pharaohs. Indeed the ancient Egyptians had a God of Perfume, Nefertem (Figure 17.1a). Nerfertem didn’t begin his mythical life as a God, but through legend and by his association with the highly aromatic, and possibly narcotic, blue water-lily flower, he rose to become the divine representation of perfume and luck. The personal wearing of scent was first recorded by the Egyptians who put flowers, herbs, and spices into wax cones that they wore on their heads; as the wax melted the aromatic mixture flowed out and perfumed them.
FIGURE 17.1
(See Color Insert)A limited pictorial survey of perfume and its vessels. (a) Nefertum, the Egyptian God of perfume and luck. (From file “Nefertem.svg” by Wikimedia Commons user Jeff Dahl. File at http://commons.wikimedia.org/wiki/File:Nefertum.svg.) (more…)
The Persian philosopher and physician Avicenna (ca. 980–1037) introduced the process of extracting oils from flowers by distillation: the method of boiling a liquid mash through which chemicals with different properties can be separated. This method is still used today.
The Etruscans revered perfume to the point that Etruscan women were never without it. The Etruscan spirit of adornment “Lassa” is a naked winged female carrying a perfume bottle. She is depicted on the engraved brass mirrors that dead Etruscan women were buried with to accompany them to the afterlife. The Romans were also great connoisseurs of perfumes and gladiators are said to have applied a different scented lotion to each area of their body before a contest. But as Christianity rose with its severe and simple attitudes towards adornment, perfume evaporated in the mist. Fortunately, this austere attitude towards self-scenting was not to be a permanent obstacle to the fragrance-seeking nose.
Perfumery came to Europe in the fourteenth century when, in 1307, at the behest of Queen Elizabeth of Hungary, the first modern perfume made of scented oils blended in an alcohol solution was produced. This perfume was thereafter referred to as “Hungary Water.” As European cities were becoming more and more fetid, the prevalence of donning perfume became ever greater. In the Renaissance perfume fashion was truly reborn and France became the epicenter of perfume development and culture, a position it has retained ever since. By the sixteenth and seventeenth centuries, the craze of perfuming everything was so extensive that even pets and jewelry were daubed with their owner’s favorite scents. And if scenting your pet sounds like an eccentricity of the past, welcome to the world of today’s perfume and pet fanatics. A spate of canine fine fragrances are currently on the market from high-end fashion designers like Juicy Couture, who offer Pawfum, at $60.00 for a 1-oz bottle of eau de parfum spray, as well as boutique perfumers, such as Renee Ryan, who in 2006 launched Sexy Beast, described as “a unisex blend of bergamot and vanilla-infused musk combined with natural patchouli, mandarin and nutmeg oils” (the 1.7-oz bottle sells for $50). For a thrifty alternate you can purchase Oh My Dog! by Etienne de Swardt, a mere $30.00 per 3.7 oz bottle (Figure 17.1b). The list goes on and cat lovers are not excluded.
By the end of the eighteenth century perfume was enjoying the status of high fashion and the higher one’s importance the better one’s fragrance. In 1709 a French perfumer proposed that the different classes should be scented differently. He concocted a royal perfume for the aristocracy, a bourgeois perfume for the middle classes, but said the poor were only deemed worthy of disinfectant. The court of Louis XV, king of France from 1715 to 1774, was known as “La Cour Parfumée” (The Perfumed Court), and the aristocracy were expected to wear a different perfume for every day of the week. So cherished were these objects of the nose that Marie Antoinette’s perfume bottle has been recently recreated, and in an extremely limited edition a 25 ml flask may be purchased for 8,000 Euros (approximately $10,290 US dollars (Figure 17.1c).
From the Renaissance into the nineteenth century, perfume wearing and perfume type were ungendered, and men and women adorned them equally. The deodorizing drive of the mid-nineteenth century, however, led to a demise of perfume and a new conservative outlook towards it. Due to the promotion of germ theory and the understanding that filth (which usually smells) carried illness, scents of all kinds began to be perceived as evil. Perfume receded to the background and took on a muted public image, and wearing fragrance became gender stereotyped. Sweet floral blends were deemed exclusively feminine, while sharper, woodsy, pine, and cedar notes were characterized as masculine. In the early to mid-twentieth century, men with any credible social position had stopped wearing fragrance altogether and were only expected to smell of clean skin and tobacco, while women of respectable social standing were expected to smell only faintly of floral notes (Classen, Howes, and Synnott 1994). Only prostitutes and the déclassé dared wear the once prestigious heavy and animalic scents of earlier generations.
A break in American perfume repression came unexpectedly during the otherwise conservative era of the 1950s. Chanel No. 5, created in 1921, was the fifth fragrance in a line developed by Ernest Beaux for Gabrielle “Coco” Chanel. Chanel No. 5 enjoyed popularity in France and Europe after its inception, but became a blockbuster when it was launched in the United States in the early 1950s, and Marilyn Monroe famously said that all she wore to bed was “two drops of Chanel No. 5.” Since the mid-1950s, Chanel No. 5 has been the most famous perfume in the world and it continues to outsell many of its modern rivals. Chanel No. 5 was also the first fragrance to be created using synthetic chemicals. Before synthetics were used, scents faded quickly and perfumes had to be continuously reapplied.
Among the classic and trendy fine fragrances of today there are also many unusual options for the perfume esthete. Among the more atypical are those available from Demeter Fragrances, which boasts over 150 scents ranging from the playful to the shocking, including: Holy Water, Dust, Playdoh, Funeral Home, Paperback, and Gin and Tonic—the art savvy can even find “This is not a Pipe.”* Demeter has not restricted itself to the entirely iconoclastic demographic and wisely has capitalized on one of the top trends in fine fragrance today—food—with eau de Birthday Cake and Sushi. Demeter isn’t the only company vying for the nose of the daring gourmand. The Stilton Cheese Makers Association, located in Surrey, England, recently launched Eau de Stilton as part of their 2006 campaign to encourage people to eat more of their cheese. The perfume, blended by a Manchester-based aromatics company, features a “symphony of natural base notes including yarrow, angelica seed, clary sage and valerian.” According to Nigel White, a company spokesman, Eau de Stilton “recreates the earthy and fruity aroma” of the cheese “in an eminently wearable perfume.” And if you want to mix the savory with sex, Burger King has the new cologne for you. Flame is, according to the company, “the scent of seduction, with a hint of flame-broiled meat.” Red meat is a favorite food among men (Drewnowski 1992; Weaver and Brittin 2001). However, if this cologne is aimed at a heterosexual male market then advertisers may have a problem. Women do not rank red meat as a highly preferred food (Drewniewski et al. 1992; Kubberod et al. 2002) and as such Flame is more likely to attract hungry men than lusty women.
On the subject of lust and women, Chanel No. 5 was not only hurtled into fame by a sex goddess but is also tainted with the odors of sex itself. Chanel No. 5 is in the scent category “floral aldehyde” and is composed of aldehydes, jasmine, rose, ylang ylang, iris, amber, and patchouli notes. But Chanel No. 5 also contains secretions from the perineal glands of the civet cat—secretions with a strong musky, fecal odor. The anal secretions from the Himalayan civet cat, musk deer, and beaver (castoreum), and vomit from sperm whales (ambergris), have been historically used as perfume fixatives. Notably, with pressure from animal rights groups, the Chanel company claimed that as of 1998 natural civet was replaced with a synthetic substitute. Many of the most popular perfumes are laden with synthetic fecal notes, such as indole. Eternity (1988) by Calvin Klein is claimed to be one of the most indolic perfumes ever. The great French perfumer Jacques Guerlain once said that perfumes should smell of “the underside of my mistress.” The scents he created, such as Jicky (1889) and Shalimar (1925), were tinged with vaginal and anal smells. Christian Dior’s perfume called Dioressence (1969) was dubbed le parfum barbare (the barbaric perfume) and smelled of animalic and fecal notes. It is an interesting social observation that within the conservative palate of the 1950s and our modern obsession with ridding the body of its own body odor, the rise of perfumes that are redolent with fecal and bodily scents re-emerged. Yet, the presence of these funky, animalic notes was rarely advertised.
We are still obsessed with bathing and eliminating our natural body odor, but curiously a new wave of fragrances are now being marketed that are deliberately aimed at recreating the scents of body funk. Secretions Magnifiques, by L’Etat Libre d’Orange, is claimed to smell like a mix of blood, sweat, saliva, and semen, and even comes in a box emblazoned with a cartoon penis squirting semen. Les Liaisons Dangereuses by Kilian is “an orgy in a perfume bottle, a fragrance steeped in the scents of group sex…” “Bodies slick with sweat, hot with the odors of sexual favors,” claims Kilian Hennessy, the perfumer who concocted it for By Kilian, his upscale, upstart Parisian perfume house. And Tom Ford, the fashion designer, reportedly told Estée Lauder executives that he wanted Black Orchid, his first fragrance, to smell like “a man’s crotch.” The irony of this new fetish for bodily and sexually scented perfumes is that our own natural body odor elicits serious responses and consequences for our sexual desire and reproductive biology, especially for women (see Section 17.4).
17.3. THE SOCIOLOGY OF PERFUME OR WHY WE WEAR PERFUME
The reasons for wearing perfume have been found to vary with psychological and demographic factors. In a study on women’s psychology of fragrance that this author conducted for the Sense of Smell Institute in 2003 (http://www.senseofsmell.org/papers/R. Herz Survey Study Final Report w. tables.doc), it was found that the socio-cultural factors that affect wearing perfume in North America are very much like those that affect clothing fashion. Young women were found to be most conformist in their perfume preferences, picking fragrances that are popular and/or that their peers are wearing. Women in their 40s were most individualistic and choose perfumes that they personally like without much consideration for outside opinion, while women aged 60 and above tended to choose perfumes that significant others had told them they enjoyed. A similar study conducted in the late 1980s for the Sense of Smell Institute showed comparable age-based trends but was a decade advanced. For example, women in their 30s were most oriented towards self-pleasing and women in their mid-40s and older were more oriented towards pleasing others (http://www.senseofsmell.org/papers/S._Schiffman_Ages_&_Stages_1991_1.doc). It is noteworthy that the differences in these data parallel the differences in the perceived role of women in society between the 1980s and the twenty-first century; 40 really is the new 30.
In Herz’s 2003 fragrance study, mood was another factor shown to influence perfume choice. Women often reported that they chose a particular scent because it had a positive effect on their mood and/or was consistent with their mood at that time. Similarly, personality influenced perfume preferences depending upon how one saw oneself (e.g., “dramatic” = heavy, oriental fragrances, or “sporty” = light, fresh fragrances). The situation or aim of the occasion (e.g., a romantic encounter or a job interview) was also a factor in what types of fragrances women selected.
Recent research has not addressed the effects of fragrance wearing on person perception. However, two studies from the 1980s suggested that female use of fragrance can elicit a negative reaction from men in a professional context (Baron 1983, 1986). The studies assessed female confederate job applicants when they wore or did not wear a popular perfume (Jontue). In both studies male subjects (interviewers) devalued the candidates’ job-related abilities when they wore fragrance, especially if it was combined with the candidate displaying other positive non-verbal cues. The women interviewers (subjects) did not show this negative bias. There are several possible reasons for this finding, which may not make it generalizable to the present. First, in the mid 1980s women had not achieved the current degree of equality and decrease in sexist valuations that they are currently (at least overtly) afforded. Secondly, although Jontue was selected as the most highly liked perfume from a pre-test sample, it may nevertheless have elicited a non-professional connotation among the male subjects, who were young (college students), presumably did not have prior experience in the real hiring process, and who may have only ever experienced this perfume in romantic settings. The context in which odors are presented in is a very powerful determinant of both their connotation and denotation (Herz and von Clef 2001; Zellner, Bartoli, and Eckard 1992). Thus, the meaning of Jontue to the male subjects may have led to a “dating” association and been incongruent with the “professional” setup. Incongruence between a fragrance and a context also results in negative evaluations (Fiore, Yah, and Yoh 2000).
As mentioned earlier, the gendering of perfume is a recent phenomenon and the socio-cultural factors that contribute to men and women’s wearing of perfume are different. Since the Renaissance, women have been wearing fragrance. By contrast, the early twentieth century saw the rejection of fragrance by men. However, by the end of the twentieth century, fragrance for men had again become a prestigious fashion statement. In the United States, men’s prestige fragrance (fragrances sold at department stores for at least $50.00) topped $900 million in 2006, with Acqua di Gio by Georgio Armani being the current number one seller (Herz 2007).
Men and women also differ in their psychological motivation for buying/wearing fragrance. Most heterosexual men principally wear fragrances that the women in their lives have given them rather than those they buy for themselves, and when men do buy fragrances they do so primarily because they believe it will attract women. By contrast, women predominantly acquire fragrance by purchasing it for a multitude of reasons, as outlined above (Herz 2007).
17.4. THE BIOLOGY OF BODY ODOR AND SEXUAL ATTRACTION
Traditional research on gender differences in human sexual behavior describes men as especially motivated by a woman’s good looks and women as most motivated by a man’s resource potential (see Buss and Schmidt 1993 for review). The explanations given for these differences are based on parental investment theory (Trivers 1972). The tenets of parental investment theory are based on the selfish gene motivation (Dawkins 1976) of striving to have the greatest representation of one’s genes in future generations—the measure of one’s reproductive success—in concert with the differential physical and behavioral costs and benefits that males and females incur in parenting.
In the cost-benefit analysis of human reproductive behavior, females invest far greater energy, resources, and time in reproduction than men, and women can only become pregnant from one man at a time (high cost). However, the benefit for women is that they are always sure of maternity; certain that the child they are investing in represents their genes and therefore facilitates their reproductive success. By contrast, the cost of reproduction for males is low but prior to the advent of genetic testing paternity was never certain. Thus, there is only a probabilistic relationship between male investment in a given child and the fact that he is investing in his own genetic material. Extrapolating from these principals, male mate selection strategies should favor being predisposed towards women who are most likely to be fertile, thus raising the probability that his genes will be passed onto future generations. By contrast, female mate selection strategies should ensure the selection of men who are most likely to secure offspring survival and thus increase the likelihood that her children (genes) will survive to be reproductively viable.
A number of cross-cultural studies have shown that males consider the female features of full lips, clear and smooth skin, clear eyes, high activity level, and a waist-hip ratio of 7:10 as attractive. These features are in fact signals of youthfulness, fertility, and potential child-bearing ability (Buss and Schmitt 1993; Hens 1995; Furnham, Tan, and McManus 1997). Thus, male mate-search strategies are predominately based on the evaluation of physical cues to fertility.
By contrast, the most adaptive strategy for females is to find males who signal that they can secure offspring survival. One way in which males can signal this ability is in the amount of material resources they can commit to a particular female and her children. A number of survey studies have indeed found that women show preferences for males with higher earning potential, ambition, and industriousness (Buss 1989; Buss and Schmitt 1993; Greenlees and McGrew 1994; Landolt, Lalumiere, and Quinsey 1995). However, far beyond the ability to take care of a woman and her children, the most important factor for a woman’s reproductive success is the likelihood that her children will be healthy enough to survive and reproduce children themselves. Therefore women should be most concerned with signals indicating a man’s physical health and most importantly signals that would indicate that the children they may conceive together will be maximally healthy.
Physical health is principally determined by one’s immune system—specifically the genes of the major histocompatibilty complex (MHC) or human leukocyte antigens (by convention the term MHC will be used here). The MHC comprises over 50 alleles and is more polymorphic in extent than any known physiological system. No two individuals other than identical twins share the same set of MHC alleles. In an ideal situation, genetic compatibility between a specific mating couple would confer MHC allele combinations to offspring that maximize disease protection from invading micro-organisms and minimize deleterious recessive mutations. Thus, MHC alleles of parents should be dissimilar; in this way the positive genes of each are most likely to be expressed and the recessive mutations least likely to be replicated. Extrapolating to behavioral strategies, females should seek males whose MHC alleles are maximally dissimilar to her own. In other words, females should be sensitive to cues that are indicative of a male’s immunological genotype.
Several studies have shown that MHC genotype influences mate choice in mice (Egid and Brown 1989; Potts, Manning, and Wakeland 1991; Yamazaki et al. 1976). Mice who are genetically identical except for minor variations in MHC loci will preferentially select mates who are dissimilar at these same loci. This discrimination has been shown to be based on the odor type of the mice (Boyse, Beauchamp, and Yamazaki 1987; Egid and Brown 1989; Yamazaki et al. 1979). Importantly, it is the female mouse which most actively makes these odor-based selections (Eklund, Egid, and Brown 1992).
Recent research among humans has also shown that MHC type plays a role in the selection of heterosexual mates. Studies on the North American Hutterite community have revealed that mate choice is influenced by an avoidance of spouses with a high degree of allele overlap with self (Ober 1999). Moreover, a number of studies by Ober and colleagues have shown that Hutterite couples with high rates of shared MHC antigens have lower fecundity and higher miscarriage rates than couples with low rates of shared MHC alleles (Ober et al. 1992, 1997, 1998). The negative consequences of MHC similar mating in the general population are further supported by fertility clinic data where higher rates of MHC similarities between couples are associated with a greater likelihood of infertility and recurrent spontaneous abortions (Ho et al. 1990; Thomas et al. 1985; Tiercy, Jeannet, and Mach 1990; Weckstein et al. 1991).
In the laboratory, Wedekind, Seebeck, Bettens, and Paepke (1995) found that females who were not on birth control pills preferred the smell of T-shirts worn by men within the test sample whose MHC genes most differed from their own. These women also reported that the scents of the T-shirts they preferred reminded them of their current and/or ex-mates. Thus, as with rodents, human mate selection appears to be influenced by female preferences for male body odors that correlate with MHC dissimilarity. As an interesting aside, one study has suggested that individuals with similar MHC types tend to share preferences for specific perfume ingredients (e.g., heliotrope) (Milinksi and Wedekind 2001). However, this finding has not been replicated or elaborated.
Recently, two survey studies found that women rank how a man smells as the most important physical trait in their choice of a sexual partner and that it is more important than all social and material status factors (Herz and Cahill 1997; Herz and Inzlicht 2002). Importantly, whether a man smelled especially attractive as a function of his natural body odor or the use of fragrance were both highly influential for female choice. That is, a man can seduce a woman on the basis of artificial fragrance and potentially mask a body odor that would indicate genetic incompatibility. This suggests that perfume can be a biological liability. Not only might an unsuitable biological match yield a less than optimally healthy child, the mismatch also increases the difficulty of conceiving (Ho et al. 1990; Thomas et al. 1985; Tiercy et al. 1990; Weckstein et al. 1991).
As professional women in the developed world postpone childbearing to their later 30s and early 40s, along with the increased infertility that comes with age, the male use of fragrance, and hence masking of incompatible genetics during courtship, may be a contributing factor to the current epidemic of infertility within this demographic. That is, women may be disproportionally selecting men who are genetically incompatible with them because they are unable to discern their “real” body odor during the initial phase of the relationship. After they discover what their man’s real body odor is like it is usually too late because of the emotional attachment now formed to him. That is, because of the facility of the olfactory system to learn the emotional significance of odors through associative learning, the scent of a genetically incompatible mate will become attractive as a function of being experienced with other rewarding aspects of the suitor (a love bond, his charm, his earning power, etc.); see Section 17.6.
17.5. PERFUME, BODY ODOR, AND THE QUESTION OF HUMAN PHEROMONES
The discussion of body odor and attraction invariably leads to the question of human pheromones. Research on this topic is currently unresolved. We do not have the organ nor corresponding neural tissue to perceive pheromones as other mammals do, and data obtained for the most substantiated human pheromone, menstrual synchrony effects (e.g., McClintock 1971; Stern and McClintock 1998), has been criticized on statistical grounds (Wilson 1992; Weller and Weller 1997). Nevertheless, several new findings suggest that we may transmit and adorn aroma-chemicals that influence our sexual motivations.
The chemical androstadienone is a steroid derivative of the male sex hormone testosterone and its presence in body fluids (e.g., sweat) is higher in males than in females. For these reasons, androstadienone has been studied as a potential human sexual pheromone. In several studies androstadienone was shown to improve women’s mood, but only when women were in the presence of men (Jacob, Hayreh, and McClintock 2001; Lundstrom and Olsson 2005). In the presence of a female experimenter, androstadienone had no impact on the participants. Another study with a male experimenter found that androstadienone increased women’s self-rated sexual arousal and cortisol levels (Wyert et al. 2007). These results have led to the speculation that androstadienone is a “modulator” pheromone for women in certain social contexts—in the presence of men. However, the levels of androstadienone that women were exposed to in these studies were a million times higher than the amount a normal male actually emits. Thus, the ecological validity of androstadienone as a human pheromone is still questionable (Herz 2008).
Notably, male use of artificial fragrances can directly augment a woman’s attraction towards him. We showed that women are more sexually responsive to a man’s scent than any other physical attribute and that this effect holds both for his true body odor and artificial fragrance (Herz and Inzlicht 2002). Moreover, recent work has shown that women visually judge men as more attractive if the men are wearing fragrance (Roberts et al. 2009). In this study, women were shown videos of men pretending to introduce themselves to an attractive woman. Half of the men in the videos had been wearing scented deodorant for two days as well as during the video taping and the other half had not. Women rated the videotapes of men who had been wearing fragrance as significantly more attractive than the fragrance-free gents, even though the women judges could not perceive any fragrance. It was further found that the men wearing the scented deodorant felt more confident than the unscented men and that the more a man liked his deodorant scent, the more confident he felt. Herz (2003) also found that 90% of all women tested in the fragrance study reported feeling more confident when they wore fragrance than when they did not (http://www.senseofsmell.org/papers/R. Herz Survey Study Final Report w. tables.doc). Thus the feelings of self-confidence inspired by wearing fragrance can alter the wearer’s behavior in a manner that increases their attractiveness to others, independent of whether those who judge them as attractive can also smell them.
With respect to chemicals emitted by women, it has been shown that the odors from breastfeeding women increased sexual desire among other women by 24% if they had a partner and 17% if they were single (Spencer et al. 2004). Thus, female body odors that vary with hormonal status may influence human sexual arousal.
Estrus is a physiological phase of the reproductive cycle of female mammals, including primates, during which there is increased female sexual receptivity, proceptivity, and attractiveness (Lange, Hartel, and Meyer 2002; Gangestad, Thornhill, and Garves-Apgar 2005). The traditional view for human reproductive biology holds that female estrus has become “lost” or “hidden” over evolutionary time (e.g., Burt 1992), presumably to promote continuous male interest and thus facilitate long-term pair-bonding and infant care-giving. However, recent studies have suggested that women during the most fertile phase of their menstrual cycle (ovulation) are most attractive to males. This increased attractiveness is manifested through superior facial attractiveness and body symmetry (Roberts et al. 2004; Manning et al. 1996), higher verbal creativity and fluency (Symonds et al. 2004), and more appealing body odor (Havlicek et al. 2006; Singh and Bronstad 2001).
Intriguingly, a recent field study on female attractiveness and hormonal status revealed that professional female lap dancers earned 80% more in tips from male patrons during ovulation than during the menses phase of their cycles (Miller, Tybur, and Jornda 2007). Thus, human females may indeed have an estrus phase and, like other mammals, are maximally attractive to men when a sexual encounter is most likely to lead to pregnancy. This finding should not be overinterpreted however, as there were no independent assessments of the dancers’ performances and therefore it is not known whether they truly did perform without variation from day to day. Female libido is known to be higher during ovulation, as are moodiness and physical discomfort during menstruation. Therefore, the dancers may not have realized that they behaved more sensually during fertile days and less so while menstruating. Moreover, what the male patrons were responding to—the dancers’ scent, looks, moves, or demeanor—is not known. At present the cause of this provocative finding is still a mystery.
17.6. THE UNIQUE CONNECTION BETWEEN OLFACTION AND EMOTION
Odors are uniquely capable of eliciting emotion. The first and most basic response we have to any scent is liking/disliking—a hedonic emotional response. Beyond simple liking, odors have the capacity to elicit full-blown emotional episodes that can be debilitating enough to require medical/ psychiatric intervention, as in the recapitulation of past trauma, and/or to bring one to overwhelming joy. Why olfaction is so potently tied to emotion is due to the mechanism by which odors come to acquire meaning, associative learning, and the neurological organization of olfaction and emotion. (For further details see Chapter 5.)
Associative learning explains both how odors become liked and disliked, as well as how odors elicit deeply emotional memory associations and behavioral responses (Bartoshuk 1991; Engen 1991; Herz 2001). After being paired with an emotionally meaningful event, a previously neutral odor can reactivate the original event, such that when later encountered the odor itself elicits the emotions that were originally paired with it, along with the consequent cognitive, behavioral, and physiological sequelae of those emotions (Herz, Beland, and Hellerstein 2004; Herz, Schankler, and Beland 2004; Herz 2007, 2009a).
Evidence supporting the associative learning hypothesis for odor perception comes from a variety of sources. First, no stereotypical responses to odors have been found in newborns, unlike the innate hedonic responses that are shown to sweet and bitter tastes. It has, however, been repeatedly shown that when presented with odorants that they have probably never encountered before, infants and children often display very different preferences from those of adults (e.g., Soussignan et al. 1997). For instance, infants find the smells of sweat and feces pleasant (Engen 1982; Stein, Ottenberg, and Roulet 1958), and toddlers do not hedonically differentiate between odorants that adults find either very unpleasant (e.g., butyric acid, found in rancid foods) or pleasant (e.g., amyl acetate, which smells like banana). Only one published study has reported that young children (three-year-olds) have adult-like responses to certain odors (Schmidt and Beauchamp 1988). However, this experiment has been criticized on methodological grounds (Engen and Engen 1997).
The olfactory system is fully functional by the third month of gestation (Schaal, Marlier, and Soussignan 1995, 1998; Winberg and Porter 1998). Therefore odor learning begins before birth, as fetuses are exposed to odorant molecules from the volatile substances that their mothers consume. This fact also supports associative learning. Mennella and colleagues found that infants of mothers who consumed distinctive-smelling volatiles (e.g., garlic, alcohol, cigarette smoke) during pregnancy or lactation showed preferences for these smells compared to infants who had not been exposed to these scents (Mennella and Beauchamp 1991, 1993; Mennella, Johnson, and Beauchamph 1995). These early learned odor preferences can continue to influence food and flavor (primarily produced by smell) preferences in later childhood (Mennella and Garcia 2000) and adulthood (Haller et al. 1999).
Associative learning can even negate flavor preference trends. In a recent study of infant formula acceptance, it was shown that when neonates were exposed to an “offensive” formula flavor, they accepted it and showed preference for this formula in later childhood. However, infants not exposed to this flavor showed negative responses to it when tested; naive adults also evaluated the flavor as unpleasant (Mennella and Beauchamp 2002). Notably feeding, in addition to providing nutrition, is an opportunity for close physical contact and emotional bonding. In both rodents and humans, association through affectionate cuddling also induces preferences for specific (yet arbitrary) scents, such as cherry oil or mother’s perfume (Balough and Porter 1986; Davis and Porter 1991; Lott, Sullivan, and McPherson 1989; Schleidt and Genzel 1990; Sullivan et al. 1991).
Cross-cultural data provides further support that associative learning, rather than hardwired responses, dictate olfactory preferences. No empirical data have shown cross-cultural consensus in odor evaluation for either common “everyday” odors (Ayabe-Kanamuura et al. 1998; Schleidt, Hold, and Attila 1981) or even “offensive” scents. Indeed, in a recent study undertaken by the U.S. military to create a “stink bomb” it was impossible to find an odor (including U.S. army issue latrine scent) that was unanimously considered unpleasant across various ethnic groups (Dilks, Dalton, and Beauchamp 1999). The following example illustrates how associated emotion is at the root of these effects.
In the mid-1960s, in Britain, Moncrieff (1966) asked adult respondents to provide hedonic ratings to a battery of common odors. A similar study was conducted in the United States in the late 1970s (Cain & Johnson, 1978). Included in both studies was the odorant methyl salicylate (wintergreen). Notably, in the British study, wintergreen was given one of the lowest pleasantness ratings, whereas in the U.S. study it was given the highest pleasantness rating. The reason for this difference can be explained by history. In Britain, the smell of wintergreen is associated with medicine and particularly for the participants in the 1966 study with analgesics that were popular during World War II, a time that these individuals would not remember fondly. Conversely, in the United States, the smell of wintergreen is exclusively a candy mint smell and one that has very positive connotations.
Empirical evidence supporting the emotional associative learning hypothesis for odor perception was shown in an experiment assessing autonomic responses to the odor of dental cement (eugenol). Patients who had previous negative experiences with dentist visits and who were fearful of dental procedures showed autonomic responses clearly indicative of fear when exposed to eugenol in the laboratory, while patients with no prior negative dental history did not (Robin et al. 1998). Most recently, laboratory studies directly aimed at testing the associative learning hypothesis for olfaction showed that a novel odorant could be made to be perceived as good or bad as a function of the emotional associations (good or bad) that were learned to it (Herz, Beland, and Hellerstein 2004).
The highly associable and emotionally evocative properties of odors are further substantiated on neuroanatomical grounds. Olfactory efferents have a uniquely direct connection with the neural substrates of emotional processing (Cahill et al. 1995; Turner, Mishkin, and Knapp 1980). Only two synapses separate the olfactory nerve from the amygdala, a structure critical for the expression and experience of emotion (Aggleton and Mishkin 1986) and human emotional memory (Cahill et al. 1995); and only three synapses separate the olfactory nerve from the hippocampus, which is critically involved in various declarative memory functions and associative learning (Eichenbaum 2001; Schwerdtfeger, Buhl, and Germroth 1990). Associative learning of specific cues (e.g., odors) to emotion is also mediated by the amygdala (LeDoux 1998) and the orbitofrontal cortex (OFC), to which olfactory processing is highly localized (see Rolls 1999). Moreover, the OFC is responsible for assigning affective valence—that is, the reward value of stimuli (Davidson, Putnam, and Larson 2000)—and fMRI experiments have found specific neural activations in the OFC for pleasant versus unpleasant odors (Gottfried et al. 2002; Anderson et al. 2003; Rolls, Kringelbach, and de Araujo 2003).
Due to the intimate neural circuitry between the olfactory cortex and the amygdala, emotional responses triggered by odors can occur instantly upon odor exposure and thus seem immediate and without conscious/cognitive mediation. Often after the emotion is experienced the associated event comes to mind, but the immediacy and priority of the emotional response makes the phenomenology of olfactory experiences different from other stimuli. In odor-evoked recall the emotional response comes first and then the formation of a cognitive association returns (though not always) later. By contrast, for all other triggers of mood and memory, the cognitive meaning precedes the assessment of emotional meaning (Herz 2007). Odors also elicit more emotional and evocative memories than any other sensory cues (see Herz 2009b for review). Indeed we have demonstrated this in an fMRI experiment concerning perfume (Herz et al. 2003).
For our study, we interviewed potential female (only) volunteers to determine whether a specific perfume could be identified that elicited a positive autobiographical memory. The criterion for participant selection was recalling a positive, personal memory in which both the smell and sight of a perfume figured. For example, one participant stated that her memory was: “A trip to Paris when I was in 4th grade and me sitting and watching my mother while she was getting ready to go out and the Opium perfume that she used which was on her vanity.” Individuals who met these criteria were invited to participate in the main experiment and one to two months later were scheduled for fMRI testing.
Testing followed a block design in which three blocks of 16 trials were administered consisting of four stimuli and clean air. Each trial lasted 30 seconds and involved the participant smelling or seeing their personally meaningful perfume (experimental odor [EO], experimental visual [EV]) and smelling or seeing a generic unmarketed perfume (control odor [CO], control visual [CV]). Each stimulus was presented twice per block, alternating with air. The order of stimulus presentation was randomly determined across blocks and subjects. The first trial of every block and between each stimulus trial was an air-only trial. During the air-only trials participants were asked to clear their mind as much as possible. During the olfactory and visual trials, participants were asked to consider whether the stimulus evoked a memory, and if so to remain thinking about that memory while the stimulus was present. After scanning, participants were presented with their EO, EV, and the CO and CV, and asked to rate the emotionality of their experience that accompanied it during the scanning procedure.
fMRI analyses revealed significantly greater activation in the amygdala and hippocampal regions during recall to the odor of the personally significant perfume (EO) than to any other stimulus. This is particularly noteworthy because odors generally elicit activation in these limbic structures. Thus the present finding was due to the distinctive emotionality of the perfume-evoked memory that was elicited and was not an olfactory artifact. Furthermore, behavioral testing confirmed that participants experienced significantly more emotion when exposed to their personally meaningful fragrance than any other stimulus (see Figure 17.2). This result is the first neurobiological demonstration that the subjective experience of the emotional potency of perfume-evoked memory is specifically correlated with heightened activation in the amygdala-hippocampal region during recall.

FIGURE 17.2
(a) Activation for the experimental odor (EO) in the amygdala. The positive activation difference for the comparison EO vs. EV+CO+CV is shown. EO = experimental odor; EV = experimental visual; CO = control odor; CV = control visual. The slice shown is (more…)
Associative learning and the immediate neural and emotional responses that odors elicit explain how some odors have earned the reputation of having “aromatherapeutic” effects, such as increasing positive mood and heart rate. Note, however, that an odor which elicits such a response is probably doing so by triggering a learned emotional association, which produces a specific mood and physiological consequence. For example, a scent connected to feeling happy and energized can indeed elicit the feelings of energy and vigor as well as heightened heart-rate or blood pressure. It should be noted that no direct pharmacological properties for odors have yet been demonstrated (Herz 2009a).
17.7. IS PERFUME DISTINCTLY HUMAN?
Are humans the only animals to adorn themselves with odor for the purposes of self-pleasure and to attract conspecifics? Many animals are known to cover themselves in scents for the purposes of defense or predation. For example, wolves will roll their bodies in carcasses or feces, presumably because this masks their own scent, which thus enables them to invisibly ambush prey. Adorning with scent therefore has adaptive purposes for other animals besides humans, but do any other animals deliberately don artificial fragrance for the purpose of pleasure?
According to researchers at the Dallas Zoo, who were trying to find scents they could use to lure ocelots into mating areas as well as move them away from dangerous areas such as highways, it was serendipitously discovered that a bottle of Obsession for Men cologne was the solution. The felines had paid no attention to various natural wild scents, but when one researcher tested her boyfriend’s bottle of Obsession, the ocelots immediately started rolling around in the scent and rubbing themselves with it. According to the zoo curator: “They tried to adorn themselves with it” (New Scientist, April 10, 1999). It was speculated that this was because of the musky indolic components in the perfume. Female ocelots in particular went wild for Obsession for Men. Male ocelots were also drawn to the scent, but for unknown reasons the male response is more variable. The use of artificial fragrance by zoo animals is not unique to ocelots, as captive gorillas have been known to rub perfume-sample cards from magazines on themselves. However, this behavior by our primate cousins may be due to the deprived context of captivity. Thus, whether we are alone or not in the personal application of fragrance for pleasure is currently unclear.
Nonetheless, the human drive towards perfume seems unparalleled. The drive is hedonic and sensual as well as psychological and manipulative. We use perfume to deliberately create pleasure and to attempt to manipulate the mood and behavior of others, especially the passion of a desired conspecific. Indeed, the holy grail of the perfume industry is to find a chemical(s) that when added to fragrance would increase the sexual interest of an intended other (Figure 17.1d).
17.8. IS PERFUME A SENSORY REWARD?
The interface between sensation and reward has been a central topic of this volume. The question therefore must be asked whether perfume is a sensory reward. On the surface the answers appears to be yes. Perfume is both intrinsically and extrinsically a sensory reward above all else. Intrinsically, perfume is created for and used for pure pleasure more than for any other function. Extrinsically, the use of perfume has rewarding properties in sexual benefits by attracting mates, and perfume may also facilitate social status and attractiveness. For example, the use of perfume can mask body-odor signals of poor health and hygiene, and because women find a man’s scent such a powerful attractant, perfuming may particularly benefit the biological fitness of men who may not otherwise be attractive.
However, as mentioned earlier, there are biological perils to perfume. Perfuming oneself, especially as a man, is not particularly beneficial for the individual or species from the point of view of attracting female mates who will be optimal biological matches and hence maximize reproductive success and child health. The benefits of female perfume use are questionable too. If females are emitting chemical signals indicative of their fertility, as possibly suggested by the lap-dancer data (Miller, Tybur, and Jornda 2007), then potential mates would be biologically served to know it.
The second assessment of the reward value of perfume is whether it serves as a reinforcer for human behavior. There have been no empirical investigations regarding the reinforcing value of perfume. However, observation of human behavior suggests that perfume is an inherently reinforcing stimulus, to the extent that it is deliberately sought out and its application is repeatedly performed with the apparent outcome of pleasure. Thus, within a classical conditioning paradigm one might predict that a favored perfume could act like an unconditioned stimulus, such that another neutral stimulus would gain reinforcement value after being paired with the perfume. This is anecdotally supported by the experience we may have upon meeting a stranger who is wearing the same perfume as someone we have very fond feelings towards—such encounters can generate a positive bias in our attitudes towards the stranger. This supposition however needs to be put to rigorous test. In all likelihood, a favored perfume initially comes to be preferred as a consequence of appetitive classical conditioning, for example, the association of a particular (hedonically inert) scent with positive emotions or with a loved one. After the perfume becomes endowed with conditioned value, it can then function as a second-order conditioned reinforcer (Holland and Rescorla 1975), conferring positive value on other stimuli through conditioned chaining. Indeed, the use of emotionally valenced odors as conditioned reinforcers has been demonstrated in both psychological (Todrank et al. 1995; Hermann et al. 2000) and neuroimaging (Gottfried, O’Doherty, and Dolan 2002, 2003; Gottfried and Dolan 2004) studies of human olfactory associative learning.
17.9. CONCLUSIONS AND THE FUTURE
It is evident that the scented human body has biological significance for mate selection and reproductive success. The chemicals in our sweat, both our unique body-odor and possibly other chemo-signals that may or may not be smelled per se, have a strong bearing on our fecundity. Additionally, adorning artificial scent increases mate attraction through various mechanisms such as increasing self-confidence or seducing a biologically incompatible mate.
What is not yet determined is whether perfume in itself is a true reward. From a neurobiological perspective, it has been shown that positive and negative odors elicit different patterns of activation (Gottfried et al. 2002; Anderson et al. 2003; Rolls, Kringelbach, and de Araujo 2003; Winston et al. 2005). Earlier work on hemispheric lateralization suggested that the left hemisphere may be dominant for positive emotional experience (Davidson 1984) and in this regard it is notable that we found a tendency for more robust activation to the personally meaningful perfume in our fMRI study in participants’ left hemispheres (Herz et al. 2003). However, a left laterality effect for personally positive odors is not entirely consistent with other neurological findings addressing odor pleasantness (Zald and Pardo 1997; Royet et al. 2003). Methodological differences in the aims of the limited studies conducted may be responsible for the lack of consistency and here is undoubtedly an area in which more work needs to be done.
Another neurological avenue for assessing perfume’s rewarding properties involves the role of the OFC. The OFC is involved in both reward and olfactory processing. However, the degree to which perfume experience specifically recruits the OFC and perhaps sub-regions thereof is not known and would be a very interesting topic to study. It is also known that the dopaminergic neurotransmitter system is highly involved in the experience of reward, and that dopaminergic pathways innervate the OFC and amygdala (Schienle et al. 2009). However, to date there are no data in humans assessing whether hedonically positive odors have a particularly activating effect on dopaminergic transmission. Future research comparing emotional and non-emotional olfactory stimuli and the neurotransmitter systems subserving them is a new topic awaiting pioneering pursuit.
From a cognitive-behavioral perspective, it has been shown that perfumes can elicit highly emotional associations and responses. However, the perfume in question must first have acquired emotional significance for the smeller through associative learning. Using behavioral paradigms in concert with neurobiological techniques, the responses elicited by a perfume before and after its association to an emotionally appetitive event could be examined. This methodology would elucidate the interplay between conscious and neurobiological mechanisms and the ways in which fragrance elicits pleasure and reward.
This chapter has reviewed various facets of the perfume experience and related topics of human chemosignal emissions in seduction, sexual advertising, and reproductive success. The present review has made it clear that although it is a tantalizing area where speculation and anecdote abound, there has been little empirical research. Most importantly, perfume has barely been studied from a neurobiological perspective or evaluated as a reinforcing stimulus. Future innovative research in perfume neuroscience and reward will be a great adornment to the field.
REFERENCES
- Aggleton J. P., Mishkin M. The amygdala: Sensory gateway to the emotions. Plutchik R., Kellerman H. Orlando: Academic Press; Emotion: Theory, Research and Experience. Vol 3: Biological Foundations of Emotion. 1986:281–99.
- Anderson A. K., Christoff K., Stappen I., Panitz D., Ghahremani D. G., Glover G., Gabrieli J. D., Sobel N. Dissociated neural representations of intensity and valence in human olfaction.
Nature Neuroscience. 2003;6:196–202.
[
PubMed: 12536208
]
- Ayabe-Kanamuura S., Schicker I., Laska M., Hudon R., Distel H., Kobabyakaw T., Saito S. Differences in perception of everyday odors: A Japanese-German cross-cultural study.
Chemical Senses. 1998;23:31–38.
[
PubMed: 9530967
]
- Balough R. D., Porter R. H. Olfactory preference resulting from mere exposure in human neonates. Infant Behavior and Development. 1986;9:395–401.
- Baron R. A. Sweet smell of success? The impact of pleasant artificial scents on evaluations of job applicants. Journal of Applied Psychology. 1983;68:709–13.
- Baron R. A. Self-presentation in job interviews: When there can be too much of a good thing. Journal of Applied Social Psychology. 1986;16:16–28.
- Bartoshuk L. M. Taste, smell and pleasure. Bolles R. C. Hillsdale, NJ: Lawrence Erlbaum; The Hedonics of Taste. 1991:15–28.
- Boyse E. A., Beauchamp G. K., Yamazaki K. The genetics of body scent. Trends in Genetics. 1987;3:97–102.
- Burr C. New York: Henry Holt; The Perfect Scent. 2007
- Burt A. Concealed ovulation and sexual signals in primates.
Folia Primatologica. 1992;58:1–6.
[
PubMed: 1618432
]
- Buss D. M. Sex differences in human mate preferences: Evolutionary hypotheses tested in 37 cultures. Behavioral and Brain Sciences. 1989;12:1–49.
- Buss D. M., Schmitt D. P. Sexual strategies theory: An evolutionary perspective on human mating.
Psychological Review. 1993;100:204–32.
[
PubMed: 8483982
]
- Cahill L., Babinsky R., Markowitsch H. J., McGaugh J. L. The amygdala and emotional memory.
Nature. 1995;377:295–96.
[
PubMed: 7566084
]
- Cain W. S., Johnson F. Jr. Lability of odor pleasantness: Influence of mere exposure.
Perception. 1978;7:459–65.
[
PubMed: 704276
]
- Classen C., Howes D., Synnott A. New York: Routledge; Aroma: The Cultural History of Smell. 1994
- Davidson R. J. Hemispheric asymmetry and emotion. Scherer K., Ekman P. Hillsdale, NJ: Erlbaum; Approaches to Emotion. 1984:39–57.
- Davidson R. J., Putnam K. M., Larson C. L. Dysfunction in the neural circuitry of emotion regulation – a possible prelude to violence.
Science. 2000;289:591–94.
[
PubMed: 10915615
]
- Davis L. B., Porter R. H. Persistent effects of early odor exposure on human neonates. Chemical Senses. 1991;16:169–74.
- Dawkins R. The Selfish Gene. New York: Oxford University Press; 1976.
- Dilks D. D., Dalton P., Beauchamp G. K. Cross-cultural variation in responses to malodors. Chemical Senses. 1999;24
- Drewnowski A., Kurth C., Holden-Wiltse J., Saari J. Food preferences in human obesity: Carbohydrate versus fats.
Appetite. 1992;18:207–21.
[
PubMed: 1510463
]
- Egid K., Brown J. L. The major histocompatibility complex and female mating preferences in mice. Animal Behaviour. 1989;38
- Eichenbaum H. The hippocampus and declarative memory: Cognitive mechanisms and neural codes.
Behavioural Brain Research. 2001;127:199–207.
[
PubMed: 11718892
]
- Eklund A., Egid K., Brown J. L. Sex differences in the use of the major histocompatibility complex for mate selection in congenic strains of mice. Doty R. L., Muller-Scwarze D. New York: Plenum Press; Chemical Signals in Vertebrates. 1992:213–17.
- Engen T. The Perception of Odors. 1982 Toronto: Academic Press.
- Engen T. Odor Sensation and Memory. New York: Praeger; 1991.
- Engen T., Engen E. A. Relationship between development of odor perception and language. Enfance. 1997;1:125–40.
- Fiore A. M., Yah X., Yoh E. Effects of product display and environmental fragrancing on approach responses and pleasurable experiences. Psychology & Marketing. 2000;17:27–54.
- Furnham A., Tan T., McManus C. Waist-to-hip ratio preferences of body shape: A replication and extension. Personality and Individual Differences. 1997;22:539–49.
- Gangestad S. W., Thornhill R., Garver-Apgar C. E. Changes in women’s sexual interests and their partners’ mate retention tactics across the menstrual cycle: Evidence for shifting conflicts of interest.
Proceedings of the Royal Society of London B. 2005;269:975–82.
[
PMC free article: PMC1690982
] [
PubMed: 12028782
]
- Gottfried J. A., Deichmann R., Winston J. S., Dolan R. J. Functional heterogeneity in human olfactory cortex: An event-related functional magnetic resonance imaging study.
Journal of Neuroscience. 2002;22:10819–28.
[
PMC free article: PMC6758422
] [
PubMed: 12486175
]
- Gottfried J. A., Dolan R. J. Human orbitofrontal cortex mediates extinction learning while accessing conditioned representations of value.
Nature Neuroscience. 2004;7:1144–52.
[
PubMed: 15361879
]
- Gottfried J. A., O’Doherty J., Dolan R. J. Appetitive and aversive olfactory learning in humans studied using event-related functional magnetic resonance imaging.
Journal of Neuroscience. 2002;22:10829–37.
[
PMC free article: PMC6758414
] [
PubMed: 12486176
]
- Gottfried J. A., O’Doherty J., Dolan R. J. Encoding predictive reward value in human amygdala and orbitofrontal cortex.
Science. 2003;301:1104–7.
[
PubMed: 12934011
]
- Greenlees I. A., McGrew W. C. Sex and age differences in preferences and tactics of mate attraction: Analysis of published advertisements. Ethology and Sociobiology. 1994;15:59–72.
- Haller R., Rummel C., Henneberg S., Pollmer U., Koster E. P. The influence of early experience with vanillin on food preference in later life.
Chemical Senses. 1999;24:465–567.
[
PubMed: 10480683
]
- Havlicek J., Dvorakova R., Baros L., Flegr J. Non-advertized does not mean concealed: Body odour changes across the human menstrual cycle. Ethology. 2006;112:81–90.
- Hens R. Waist-to-hip ratio and attractiveness: Replication and extension. Personality and Individual Differences. 1995;19:479–88.
- Hermann C., Ziegler S., Birbaumer N., Flor H. Pavlovian aversive and appetitive odor conditioning in humans: Subjective, peripheral, and electrocortical changes.
Experimental Brain Research. 2000;132:203–15.
[
PubMed: 10853945
]
- Herz R. S. Ah, sweet skunk: Why we like or dislike what we smell. Cerebrum. 2001;34:31–47.
- Herz R. S. Survey Study Final Report. 2003 http://www.senseofsmell.org/papers/R. Herz w. tables.doc.
- Herz R. S. The unique interaction between language and olfactory perception and cognition. New York: Nova Science; Trends in Experimental Psychology Research. 2005:91–109.
- Herz R. S. New York: William Morrow/ HarperCollins; The Scent of Desire: Discovering Our Enigmatic Sense of Smell. 2007
- Herz R. S. Olfaction. Sunderland, MA: Sinauer Associates; Sensation & Perception. (2nd) 2008:330–58.
- Herz R. S. Aromatherapy facts and fictions: A scientific analysis of olfactory effects on mood, physiology and behavior.
International Journal of Neuroscience. 2009a;119:263–90.
[
PubMed: 19125379
]
- Herz R. S. Basic processes in human olfactory cognition: Current questions and future directions.
Annals of the New York Academy of Sciences. 2009b;1170:313–17.
[
PubMed: 19686153
]
- Herz R. S., Beland S. L., Hellerstein M. Changing odor hedonic perception through emotional associations in humans. International Journal of Comparative Psychology. 2004;17:315–39.
- Herz R. S., Cahill E. D. Differential use of sensory information in sexual behavior as a function of gender.
Human Nature. 1997;8:275–86.
[
PubMed: 26196967
]
- Herz R. S., Eliassen J. C., Beland S. L., Souza T. Neuroimaging evidence for the emotional potency of odor-evoked memory.
Neuropsychologia. 2004;42:371–78.
[
PubMed: 14670575
]
- Herz R. S., Inzlicht M. Gender differences in response to physical and social signals involved in human mate selection: The importance of smell for women. Evolution and Human Behavior. 2002;23:359–64.
- Herz R. S., Schankler C., Beland S. Olfaction, emotion and associative learning: Effects on motivated behavior. Motivation and Emotion. 2004;28:363–83.
- Herz R. S., von Clef J. The influence of verbal labeling on the perception of odors: Evidence for olfactory illusions?
Perception. 2001;30:381–91.
[
PubMed: 11374206
]
- Ho H. N., Gill T. J., Nsieh R. P., Hsieh H. J., Lee T. Y. Sharing of human leukocyte antigens in primary and secondary recurrent spontaneous abortions.
American Journal of Obstetrics and Gynecology. 1990;163:178–88.
[
PubMed: 2165353
]
- Holland P. C., Rescorla R. A. Second-order conditioning with food unconditioned stimulus.
Journal of Comparative and Physiological Psychology. 1975;88:459–67.
[
PubMed: 1120816
]
- Jacob S., Hayreh D. J., McClintock M. K. Context-dependent effects of steroid chemosignals on human physiology and mood.
Physiology and Behavior. 2001;74:15–27.
[
PubMed: 11564447
]
- Kubberod E., Ueland O., Rodbotten M., Westad F., Risvik E. Gender specific preferences and attitudes towards meat. Food Quality and Preference. 2002;13:285–94.
- Landolt M. A., Lalumiere M. L., Quinsey V. L. Sex differences in intrasex variations in human mating tactics: An evolutionary approach. Ethology & Sociobiology. 1995;16:3–23.
- Lange I. G., Hartel A., Meyer H. H. D. Evoluation of oestrogen functions in vertebrates.
Journal of Steroid Biochmiestry and Molecular Biology. 2002;83:219–26.
[
PubMed: 12650719
]
- LeDoux J. Fear and the brain: Where have we been, and where are we going?
Biological Psychiatry. 1998;44:1229–38.
[
PubMed: 9861466
]
- Lorig T. On the similarity of odor and language perception.
Neuroscience and Biobehavioral Reviews. 1999;23:391–98.
[
PubMed: 9989426
]
- Lott I. T., Sullivan R. M., McPherson D. Associative olfactory learning occurs in the neonate. Neurology. 1989;39
- Lundstrom J. N., Olsson M. J. Subthreshold amounts of social odorant affect mood, but not behavior, in heterosexual women when tested by a male, but not a female experimenter.
Biological Psychology. 2005;70:197–204.
[
PubMed: 16242537
]
- Manning J. T., Scutt D., Whiothosue G. H., Kinnster S. J., Walton J. M. Asymmetry and the menstrual cycle in women. Ethology and Sociobiology. 1996;17:129–43.
- McClintock M. K. Menstrual synchrony and suppression.
Nature. 1971;229:244–45.
[
PubMed: 4994256
]
- Mennella J. A., Beauchamp G. K. The transfer of alcohol to human milk: Effects on flavor and the infant’s behavior.
New England Journal of Medicine. 1991;325:981–85.
[
PubMed: 1886634
]
- Mennella J. A., Beauchamp G. K. The effects of repeated exposure to garlic-flavored milk on the nursling’s behavior.
Pediatric Research. 1993;34:805–8.
[
PubMed: 8108198
]
- Mennella J. A., Garcia P. L. Children’s hedonic response to the smell of alcohol: Effects of parental drinking habits.
Alcoholism: Clinical and Experimental Research. 2000;24:1167–71.
[
PubMed: 10968653
]
- Mennella J. A., Johnson A., Beauchamp G. K. Garlic ingestion by pregnant women alters the odor of amniotic fluid.
Chemical Senses. 1995;20:207–9.
[
PubMed: 7583013
]
- Milinski M., Wedekind C. Evidence for MHC-correlated perfume preferences in humans. Behavioral Ecology. 2001;12:140–49.
- Miller G., Tybur J. M., Jornda B. D. Ovulatory cycle effects on tip earnings by lap dancers: Economic evidence for human estrus? Evolution and Human Behavior. 2007;28:375–81.
- Moncreiff R. W. New York: Wiley; Odour Preferences. 1966
- Moran J. La Quinta, CA: Crescent House Publishing; Fabulous Fragrances II. 2000
- Ober C. Studies of HLA, fertility and mate choice in a human isolate.
Human Reproduction Update. 1999;5:103–7.
[
PubMed: 10336015
]
- Ober C., Hyslop T., Elias S., Weitkamp L. R., Hauck W. W. Human leucocyte antigen matching and fetal loss: Results of a 10 year prospective study.
Human Reproduction. 1998;13:33–38.
[
PubMed: 9512225
]
- Potts W. K., Manning C. J., Wakeland E. K. Mating patterns in seminatural populations of mice influenced by MHC genotype.
Nature. 1991;352:619–21.
[
PubMed: 1865924
]
- Roberts S. C., Havlicek J., Flegr J., Hrruskova M., Little A. C., Jones B. C., et al. Female facial attractiveness increases during the fertile phase of the menstrual cycle.
Proceedings of the Royal Society of London Series B. 2004;271S5:S270–72.
[
PMC free article: PMC1810066
] [
PubMed: 15503991
]
- Roberts S. C., Little A. C., Lyndon A., Roberts J., Havlicek J., Wright R. I. Manipulation of body odour alters men’s self-confidence and judgments of their visual attractiveness by women.
International Journal of Cosmetic Science. 2009;31:47–54.
[
PubMed: 19134127
]
- Robin O., Alaoui-Ismaili O., Dittmar A., Vernet-Mauri E. Emotional responses evoked by dental odors: An evaluation from autonomic parameters.
Journal of Dental Research. 1998;77:1638–46.
[
PubMed: 9719038
]
- Rolls E. T. Oxford; Oxford University Press: The Brain and Emotion. 1999
- Rolls E. T., Kringelbach M. L., de Araujo I. E. Different representations of pleasant and unpleasant odours in the human brain.
European Journal of Neuroscience. 2003;18:695–703.
[
PubMed: 12911766
]
- Royet J. -P., Plailly J. Lateralization of olfactory processes.
Chemical Senses. 2004;29:731–45.
[
PubMed: 15466819
]
- Royet J. P., Plailly J., Delon–Martin C., Kareken D. A., Segebarth C. fMRI of emotional responses to odors: Influence of hedonic valence and judgment, handedness, and gender.
Neuroimage. 2003;20:713–28.
[
PubMed: 14568446
]
- Schaal B., Marlier L., Soussignan R. Responsiveness to the odor of amniotic fluid in the human neonate.
Biology of the Neonate. 1995;671:397–406.
[
PubMed: 7578623
]
- Schaal B., Marlier L., Soussignan R. Olfactory function in the human fetus: Evidence from selective neonatal responsiveness to the odor of amniotic fluid.
Behavioral Neuroscience. 1998;112:1438–49.
[
PubMed: 9926826
]
- Schienle A., Schafer A., Hermann A., Vaitl D. Binge-eating disorder: Reward sensitivity and brain activation to images of food.
Biological Psychiatry. 2009;65:654–61.
[
PubMed: 18996508
]
- Schiffman S. S. 1991. http://www.senseofsmell.org/papers/S._Schiffman_Ages_&_Stages_1991_1.doc.
- Schleidt M., Genzel C. The significance of mother’s perfume for infants in the first weeks of their life. Ethology and Sociobiology. 1990;11:145–54.
- Schleidt M., Hold B., Attila G. A cross-cultural study on the attitude towards personal odors.
Journal of Chemical Ecology. 1981;7:19–31.
[
PubMed: 24420424
]
- Schmidt H. J., Beauchamp G. K. Adult-like odor preferences and aversions in three year old children.
Child Development. 1988;59:1136–43.
[
PubMed: 3168621
]
- Schwerdtfeger W. K., Buhl E. H., Germroth P. Disynaptic olfactory input to the hippocampus mediated by stellate cells in the entorhinal cortex.
Journal of Comparative Neurology. 1990;292:163–77.
[
PubMed: 2319007
]
- Soussignan R., Schaal B., Marlier L., Jiang T. Facial and autonomic responses to biological and artificial olfactory stimuli in human neonates: Re-examining early hedonic discrimination of odors.
Physiology & Behavior. 1997;62:745–58.
[
PubMed: 9284493
]
- Spencer N. A., McClintock M. K., Sellergren S. A., Bullivant S., Jacob S., Mennella J. A. Social chemosignals from breastfeeding women increase sexual motivation.
Human Behavior. 2004;46:362–70.
[
PubMed: 15325237
]
- Stein M., Ottenberg M. D., Roulet N. A study of the development of olfactory preferences.
Archives of Neurological Psychiatry. 1958;80:264–66.
[
PubMed: 13558783
]
- Stern K., McClintock M. K. Regulation of ovulation by human pheromones.
Nature. 1998;392:177–79.
[
PubMed: 9515961
]
- Sullivan R. M., Taborsky-Barba S., Mendoza R., Itano A., Leon M., Cotman C., Payne T. F., Lott T. Olfactory classical conditioning in neonates.
Pediatrics. 1991;87:511–18.
[
PMC free article: PMC1952659
] [
PubMed: 2011429
]
- Symonds C. S., Gallagher P., Thompson J. M., Young A. H. Effects of the menstrual cycle on mood, neurocognitive and neuroendocrine function in healthy premenopausal women.
Psychological Medicine. 2004;34:93–102.
[
PubMed: 14971630
]
- Thomas M. L., Harger J. H., Wagner D. K., Rabin B. S., Gill T. J. III. HLA sharing and spontaneous abortion in humans.
American Journal of Obstetrics and Gynecology. 1985;151:1053–58.
[
PubMed: 3157320
]
- Tiercy J. M., Jeannet M., Mach B. A new approach for the analysis of HLA class II polymorphism: HLA oligotyping.
Blood Review. 1990;4:9–15.
[
PubMed: 2108745
]
- Todrank J., Byrnes D., Wrzesniewski A., Rozin P. Odors can change preferences for people in photographs: A cross-modal evaluative conditioning study with olfactory USs and visual CSs. Learning and Motivation. 1995;26:116–40.
- Trivers R. Parental investment and sexual selection. Campbell B. Chicago: Aldine-Atherton; Sexual Selection and the Descent of Man. 1972
- Turner B. H., Mishkin M., Knapp M. Organization of the amygdalopetal projections from modality-specific cortical association areas in the monkey.
Journal of Comparative Neurology. 1980;191:515–43.
[
PubMed: 7419732
]
- Walla P., Hufnagl B., Lehern J., Mayer D., Lindinger G., Imhof H., Deeke L., Lang W. Olfaction and depth of word processing: A magnetoencephalographic study.
Neuroimage. 2003;18:104–16.
[
PubMed: 12507448
]
- Weckstein L. N., Patrizio P., Balmaceda J. P., Asch R. H., Branch D. W. Human leukocyte antigen compatibility and failure to achieve a viable pregnancy with assisted reproductive technology.
Acta European Fertility. 1991;22:103–7.
[
PubMed: 1746211
]
- Wedekind C., Seebeck T., Bettens F., Paepke A. J. MHC-dependent mate preference in humans.
Proceedings of the Royal Society of London B. 1995;260:245–49.
[
PubMed: 7630893
]
- Weaver M. R., Brittin H. C. Food preferences of men and women by sensory evaluation versus questionnaire. Family and Consumer Sciences Research Journal. 2001;29:288–301.
- Weller L., Weller A. Menstrual variability and the measurement of menstrual synchrony.
Psychoneuroendocrinology. 1997;22:115–28.
[
PubMed: 9149333
]
- Wilson H. C. A critical review of menstrual synchrony research.
Psychoneuroendocrinology. 1992;17:565–91.
[
PubMed: 1287678
]
- Winberg J., Porter R. H. Olfaction and human neonatal behaviour: Clinical implications.
Acta Paediatrica. 1998;87:6–10.
[
PubMed: 9510439
]
- Winston J. S., Gottfried J. A., Kilner J. M., Dolan R. J. Integrated neural representations of odor intensity and affective valence in human amygdala.
Journal of Neuroscience. 2005;25:8903–7.
[
PMC free article: PMC6725588
] [
PubMed: 16192380
]
- Wyert C., Webster W. W., Chen J. H., Wilson S. R., McClary A., Khan R. M., Sobel N. Smelling a single component of male sweat alters levels of cortisol in women.
Journal of Neuroscience. 2007;27:1261–65.
[
PMC free article: PMC6673596
] [
PubMed: 17287500
]
- Yamazaki K., Boyse E. A., Mike V., Thaler H. T., Mathieson B. J., Abbot J., Boyse J., Zayas Z. A., Thomas L. Control of mating preferences in mice by genes in the major histocompatibility complex.
Journal of Experimental Medicine. 1976;144:1324–35.
[
PMC free article: PMC2190468
] [
PubMed: 1032893
]
- Yamazaki K., Yamaguchi M., Baranoski L., Bard J., Boyse E. A., Thomas L. Recognition among mice. Evidence from the use of a Y-maze differentially scented by congenic mice of different major histocompatibility types.
Journal of Experimental Medicine. 1979;150:755–76.
[
PMC free article: PMC2185685
] [
PubMed: 512584
]
- Zald D. H., Pardo J. V. Emotion, olfaction, and the human amygdala: Amygdala activation during aversive olfactory stimulation.
Proceedings of the National Academy of Sciences USA. 1997;948:4119–24.
[
PMC free article: PMC20578
] [
PubMed: 9108115
]
- Zellner D. A., Bartoli A. M., Eckard R. Influence of color on odor identification and liking ratings.
American Journal of Psychology. 1991;104:547–61.
[
PubMed: 1793126
]
*
Allusion to the surrealist artist Rene Magritte.
Other posts : [wpb-random-posts]



